

Chelicerata is a major subphylum of Arthropoda that includes horseshoe crabs and arachnids, such as harvestmen, scorpions, spiders, ticks, and mites. The chelicerates derive their name from ‘chelicerae,’ specialized appendages found anterior to the mouth that constitute an integral part of their feeding apparatus.
Like other arthropods, chelicerates have characteristically segmented bodies and jointed appendages covered with a thick, chitinous cuticle. They are one of the most diverse groups of animals, with over 77,000 extant species, including around 50,000 species of spiders, 32,000 species of ticks and mites, and 1,400 scorpion species[1].
The body of a chelicerate is covered with a thick cuticle and divided into two tagmata – the anterior prosoma or cephalothorax and the posterior opisthosoma or abdomen[2]. However, there are no visible tagmata in mites and ticks.
The prosoma forms in the embryo by fusing the ocular somite (formerly called the ‘acron,’ which lies just before the first true segment and bears the eyes and labrum) with six post-ocular segments (somites 1–6). Each of these six segments carries a paired appendage. In most chelicerates, the opisthosoma consists of up to thirteen segments, which may or may not terminate in a telson. In some taxa, such as scorpions and eurypterids, the opisthosoma is divided into two regions: a front mesosoma and a rear metasoma, with the metasoma typically lacking appendages. In modern chelicerates, abdominal appendages are either absent or highly modified. For example, in spiders, the remaining appendages form spinnerets for silk production, while in horseshoe crabs, they develop into gills[3].
Though it was initially believed that chelicerates had lost their first antennae-bearing somite, current evidence suggests that it is retained and is similar to the pair of chelicerae or chelifores. The second somite is home to the pair of pedipalps (used for sensory purposes), while the subsequent four segments (somites 3 to 6) possess pairs of walking legs. The mouth is situated between somites 1 and 2 (chelicerae and pedipalps, respectively)[4].
In most chelicerates, the chelicerae are usually divided into three distinct segments, with the third segment resembling a claw. However, spiders have only two segments; the second segment usually folds under the first when not in use. The sizes of chelicerae vary considerably, ranging from large claw-like structures in fossil eurypterids and modern harvestmen to small toothed appendages in scorpions.
The pedipalps usually perform specialized sensory functions in most chelicerate groups; however, in more derived ones, like spiders and scorpions, they are modified to pincers for catching prey and may also contain bulbous tips for injecting sperm into the females’ gonopores.
However, the chelicerae and pedipalps differ in embryonic origin. Although the deutocerebrum innervates both chelicerae and pedipalps, the former arise from somite 1, positioned ahead of the mouth, while the latter develop from somite 2, located posterior to the mouth.
In chelicerates, the cuticle is deeply innervated by multiple sensors from the nervous system. Spiders and other arthropods have evolved intricate sensory arrays from their cuticles, including touch and vibration sensors, mainly ‘setae’ (bristles), which detect a range of forces from strong contact to gentle air currents.
The primary body cavity in chelicerates is the hemocoel, which extends along most of the body and channels circulating blood. This circulation is facilitated by a tubular heart that collects blood from the rear parts of the body and propels it forward. Like other arthropods, chelicerates have an open circulatory system in which their arteries end openly, rather than connecting directly to veins.
Modern terrestrial chelicerates generally possess both book lungs and tracheae for respiration. Book lungs facilitate the exchange of oxygen and removal of waste gases through the bloodstream, while tracheae perform the same function independently of the blood transport system.
However, the aquatic horseshoe crabs respire through a horizontally placed series of thin, membranous plates called book gills. Scientists have long speculated that extinct eurypterids also possessed gills, but the fossil records were always inconclusive. Later, the discovery of the fossil of an extinct eurypterid, Onychopterella, belonging to the Late Ordovician period, confirmed the existence of four pairs of vertically oriented book gills in them.
Chelicerates are generally ammonotelic, expelling nitrogenous waste as ammonia, which is primarily released through the gills or other body surfaces. Solid waste is excreted separately as feces via the anus, after being filtered by Malpighian tubules and other excretory glands. For example, horseshoe crabs use specialized excretory organs like nephridia (also known as ‘little kidneys’) to expel ammonia.
Since ammonia demands high quantities of water for dilution, most terrestrial chelicerates, which need to conserve water, often convert nitrogenous waste into other chemicals and excrete them as dry matter.
Chelicerates have a nervous system similar to all other arthropods, consisting of a pair of nerve cords with a ganglion per segment. These nerve cords converge, forming the brain through the fusion of ganglia located just behind the mouth with pre-oral ganglia situated ahead of the mouth. Scientists previously assumed that the first segment, homologous to the antennal segment in other arthropods, was missing in chelicerates, leading to the interpretation that their brain contained only one pair of pre-oral ganglia. However, further evidence negates this assumption and reveals that the first segment, which bears the chelicerae, is present.
In some chelicerates like horseshoe crabs, the brain integrates ganglia from the prosoma and the first two opisthosomal segments, while the remaining opisthosomal segments contain separate ganglia.
Except for scorpions, in most arachnids, the ganglia fuse into a single mass in the prosoma, and the opisthosoma does not contain any such neural connection. However, Mesothelae spiders retain primitive ganglia in both the opisthosoma and the rear portion of the prosoma.
Chelicerates may have compound eyes, simple eyes (pigment-cup ocelli), or both (as in the case of horseshoe crabs). Among modern chelicerates, true compound eyes are found only in horseshoe crabs. In other groups, the eyes typically consist of clusters of simple eyes, with up to five pairs of ocelli. While compound eyes are found on both sides of the head, the pigment-cup ocelli are located either centrally or laterally, depending on the species. These median ocelli-type eyes are thought to be evolutionarily related to the nauplius eyes of crustaceans and the ocelli of insects.
There is considerable variability in the extent of visual perception in different species of chelicerates. The compound eyes in horseshoe crabs are sensitive enough to detect movement but fail to form images, whereas jumping spiders have a wide field of vision, color acuity, and UV light perception.
The traditional family tree of Chelicerata showed that these arthropods are more distantly related to the other major groups, like crustaceans, hexapods, and myriapods, than those groups are to each other. However, recent advancements in molecular phylogenetics and embryological studies on the development of arthropod nervous systems reveal a revised phylogeny. They suggest a closer relationship between chelicerates and myriapods, while the hexapods and crustaceans are reported to be the closest to each other. Although sea spiders (Pycnogonida) are also classified under Chelicerata, some features suggest they may represent the earliest arthropods from which other chelicerates evolved.
This subphylum is thought to have diverged from the mandibulate lineage of arthropods during the middle Cambrian period, although the exact period of divergence is still unknown. This division is supported by the presence of so-called early chelicerate ancestors during that period, including those belonging to the order Habeliida and the extinct genus Mollisonia.
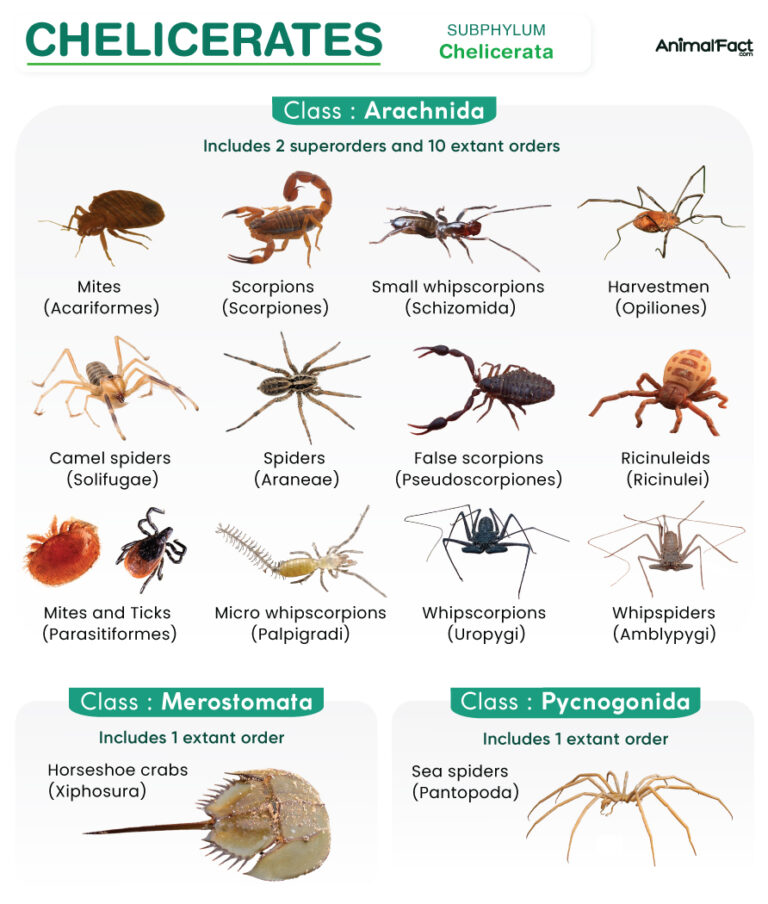
Although scientists have reached a global consensus suggesting Chelicerata include the classes Arachnida and Merostomata (Xiphosura and the extinct Eurypterida), the inclusion of Pycnogonida (sea spiders) under this subphylum remains debatable[5][6].
Based on the current classification, chelicerates are divided into 3 classes, 2 superorders, and 18 orders.
They inhabit a wide range of terrestrial, freshwater, and marine habitats worldwide.
Spiders and scorpions are terrestrial, inhabiting forests, grasslands, deserts, and urban environments, whereas water mites occupy freshwater habitats. In contrast, sea spiders and horseshoe crabs are completely marine, ranging from shallow waters to the deep sea.
Originally predators, chelicerates have diversified to use all the major feeding strategies, including predation, parasitism, herbivory, and scavenging.
Although most chelicerates are carnivores, feeding mainly on soft-bodied invertebrates, the spider Bagheera kiplingi, with a herbivorous diet, is an exception, while many others supplement their diets with nectar and pollen. Most ticks and mites are parasitic and suck on host blood, whereas harvestmen and a few other chelicerates are scavengers and detritivores feeding on solid food like detritus.
The reproductive strategies of chelicerates depend largely on the habitat in which they live.
Horseshoe crabs reproduce through external fertilization, where sperm and eggs unite outside the parents’ bodies. Despite living in water, they spawn on land in the intertidal zone of beaches. The female digs a depression in the wet sand to deposit her eggs, and then the male (often multiple males) releases sperm onto them. Horseshoe crab larvae resemble miniature adults with full sets of appendages and eyes. Initially, they bear two pairs of book gills and gain three more pairs as they molt. Sea spiders also have a similar fertilization strategy in which sperm and eggs are released into the water to be fertilized.
Most arachnids (being air-breathing) employ indirect internal fertilization, though many mites have become secondarily aquatic. In one method, the male deposits a spermatophore (a packet of sperm) on the ground, which the female picks up. Alternatively, the male may store sperm in specialized appendages, such as the pedipalps in male spiders, and transfer them to the female during mating. Courtship rituals are common, especially in species at high risk of being eaten before mating. Although most arachnids lay eggs (oviparous), all scorpions and a few species of mites are viviparous, giving birth to young ones.
Their parental care varies from a short period to a prolonged one, depending on the chelicerate species. While scorpions carry the young ones on their backs till their first molt, a few spider species, like wolf spiders, have developed specialized, rough bristles to help their young cling to their backs. Some female spiders also regurgitate food in response to the begging of their young ones.
Chelicerates are preyed upon by natural predators like birds, mammals, amphibians, reptiles, and other arthropods.
Welcome to AnimalFact - your go-to source for fascinating insights into the amazing world of animals!


